Cerebral organization of adaptive linguistic behavior in developmental stuttering
Pathophysiology of stuttering remains an object of study in modern neuroscience. Over years of research, numerous theories regarding its etiology and pathogenesis have been formulated. One of the first formal theories was the idea of incomplete or abnormal cerebral dominance, proposed by Orton [1927] and Travis [1931]. They suggested that unformed or disturbed speech cerebral dominance in people who stutter leads to a reduction of motor impulses to bilaterally innervated speech muscles. Since 1966, when Jones, using the Wada test, established bilateral speech representation in 4 stutterers, new evidence of an abnormal lateralization pattern in this category of patients has emerged. The majority of results confirmed that stutterers demonstrate differences in cerebral dominance compared to the norm [Fox et al., 1996; Ingham et al., 1994; Ingham et al., 1996; Wu et al., 1995]. Based on the results of these studies, different assumptions about the localization of speech were made: left-hemisphere dominance, right-hemisphere dominance, right-hemisphere interference with the left hemisphere, inversion of speech dominance, bilateral speech representation.
In recent years, positron emission tomography (PET) and magnetic resonance imaging (MRI) have opened new possibilities for studying the morphofunctional substrate of speech, both in normal conditions and in stuttering. These studies have refined existing concepts regarding the localization of sensorimotor and cognitive-linguistic components of speech. Normally, planned and prepared speech without a semantic component activates the primary motor area - Brodmann area 4 (hereinafter fields according to Brodmann) bilaterally [3, 5, 7, 11]. The right and left primary motor areas are symmetrically activated because the articulatory, respiratory muscles, as well as the pharynx and larynx, are bilaterally innervated. Only on the left during speech production is the secondary motor area of the cortex (area 6) activated [2, 4, 5]. Regional cerebral blood flow in the dorsolateral prefrontal cortex (areas 8, 9), medial orbital (area 11), and medial prefrontal (area 10) regions, presumably associated with semantic components of speech activity, normally increases only on the left [1, 2]. The primary auditory and auditory association cortex areas in the superior temporal gyrus (areas 41, 42, 22) are normally activated bilaterally. After primary and secondary processing in these areas, information is transmitted to frontal, temporoparietal, paralimbic, and subcortical regions [Pandya & Yeterian, 1985]. Thus, these cortical areas are associated not only with auditory perception but also with the preparation of expressive speech [2, 8, 11]. The superior opercular areas (areas 44, 45), which classically constitute Broca's area, are traditionally considered involved in speech motor programming [Freedman et al., 1984] and demonstrate unilateral left-sided activation in the norm. The anteroventral areas of the frontal operculum (area 47), activated on the left, are more closely related to syntactic processes [McCarthy et al., 1993] and utterance formation [1, 4, 11]. Only in the left hemisphere during linguistic processes are the anterior temporal pole (area 38) and the angular gyrus (area 39) activated [2, 11]. John Van Borsel and colleagues discovered the involvement of tertiary fields in the speech process — motor (area 46 bilaterally), visual (area 19 on the left, area 7 on the right), and sensory (area 40 bilaterally) [11].
In addition to cortical areas, the cerebellum is also involved in speech production in healthy right-handed subjects. It is believed that this structure is connected to the auditory cortex and contains neurons that respond to auditory stimuli [Snider et al., Stowell, 1944; Hampson 1949; Aitkin & Boyd, 1975; Huang & Liu, 1985]. The cerebellar vermis is positioned to control speech production and modulate speech motor activity via ascending efferents. Thus, the cerebellum participates in speech control and is specifically involved in detecting errors and correcting motor activity initiated by neocortical systems [Thach et al., 1992].
The insula is normally activated by acoustic stimulation [Kushner et al., 1987], and selective damage to this area can lead to a relatively specific impairment of auditory signal processing [Fifer, 1993]. In the 1970s, Mohr conducted a thorough comparative clinical and postmortem study of patients' brains and established that infarcts limited to the area of the left anterior insula and pars opercularis lead to temporary severe dysarthria or near-muteness without accompanying aphasia [Mohr et al., 1978]. According to some data, the insula may be involved in relatively direct processing of auditory information, according to others — in speech articulatory planning [Pandya et al., 1969; Mesulam & Mufson, 1982].
According to PET studies, brain metabolism in stutterers and non-stutterers at rest does not differ [Braun et al., 1997; Ingham et al., 1996]. Articulatory and pharyngeal movements during speech production without a linguistic component cause bilateral activation in the primary motor, premotor, somatosensory, and auditory cortex, insula, cerebellum, putamen, ventral thalamus, both in normal conditions and in stuttering. While there are no qualitative differences in the nature and anatomical distribution of activation, the magnitude of the increase in regional blood flow in these areas during speech production is significantly greater in stutterers [2, 11]. In the neocortex, activation is greater in the premotor, primary motor, and somatosensory cortex, meaning these areas may be more active in the generation and proprioceptive or tactile perception of movements of the lips, tongue, jaw, and larynx. Many researchers have obtained evidence of hyperactivity of the right primary motor area (area 4) in stutterers during speech compared to the norm [2, 11, 12]. The medial part of area 6 (supplementary motor area), presumably associated with speech breathing, is restricted to the left hemisphere both in the norm and in stuttering, whereas the lateral part of area 6, possibly associated with motor and phonemic planning, is activated bilaterally in stuttering with reduced activation on the left [2, 4, 5].
The cerebral organization of the linguistic components of speech, unlike the sensorimotor components, is fundamentally different in adults who stutter. Normally, during the formulation of a speech utterance, cerebral blood flow increases mainly in the left hemisphere. In stutterers, the increase in blood flow is absent, spatially restricted, bilateral, or lateralized to the right hemisphere [2, 3, 6, 9, 11, 12]. Secondary motor (areas 8, 9), medial orbital (area 11), and prefrontal (area 10) areas demonstrate unilateral left-sided activation in the norm and weak bilateral activation in stutterers [Chang et al., 2009; Wood et al., 1980; Wu et al., 1995; Fox et al., 2000]. The primary and secondary auditory fields (areas 41, 42, 22), normally activated bilaterally, show significantly lower left-sided blood flow in the primary auditory and auditory association cortex in stutterers, even at rest [Pool et al., 1991]; however, the right-hemisphere homologue of Wernicke's area (area 22) is more active [3, 6, 7, 9, 12]. The anterior temporal pole (area 38), activated during linguistic processes on the left in the norm, is activated on the right in stutterers. Furthermore, stutterers do not demonstrate activation of the left temporoparietal regions bordering Wernicke's area — the posterior superior temporal (area 40) and inferior angular (area 39) gyri [2, 11]. Activation of the right homologue of Broca's area (areas 44, 45) is observed only in stutterers [1, 2].
During speech, stutterers show significantly increased blood flow in the cerebellum bilaterally, in the insula bilaterally with spread to the putamen, and in the midbrain (substantia nigra, pedunculopontine nucleus, red nucleus, and subthalamic nucleus) bilaterally [12]. Activation of the anterior insula on the right, putamen on the right, and mesencephalic periaqueductal gray matter is observed only in stutterers [2, 3, 6, 7, 9, 11].
To this day, it has not been determined whether the specific pattern of brain activation in stuttering is the cause of the defect, a reflection of compensatory activity of the nervous system, or a consequence of adaptive behavior.
Adults who stutter report that from childhood or adolescence they control their speech to mask the manifestations of the defect and attempt to prevent occurring disfluencies using various techniques and tricks. The most common speech tricks include: substituting words for those that do not contain certain "difficult" sounds; periodic stops within words and between words to eliminate the influence of involuntary contractions of articulatory, vocal, and pharyngeal muscles on speech; frequent use before difficult words of standard, familiar, and easy function words that carry no semantic load ("well", "this", "here", etc.). Apparently, a stutterer during speech must preliminarily analyze the upcoming utterance and, if it contains words beginning with sounds that are problematic for him/her, preface them with function words or replace them with synonyms; pause during involuntary muscle contractions. Accordingly, the linguistic activity of a stutterer from childhood must be aimed not only at constructing a competent and extensive speech utterance that most accurately reflects his/her thoughts and feelings but also at minimizing the manifestations of the defect in speech. If we assume that the linguistic processes of a stutterer are simultaneously aimed both at forming the upcoming utterance and scanning the future sentence, then, in that case, stutterers, compared to fluent speakers, should perform better on speech tasks related to avoiding certain words and sounds. To test this assumption, we asked 2 groups of subjects to retell a well-known text, avoiding words beginning with specific letters.
Methods. The study involved 10 stutterers and 10 non-stutterers, all right-handed. The gender composition (8m/2f), mean age (45±9 years), and education level (12±4 years of schooling) were roughly equal in both groups. Each subject had to read a text consisting of 1000 words. After that, the subject was asked to retell it without using words beginning with the letter "n" (the text contained 45 words starting with the letter "n"). During the 3 minutes of retelling, the experimenter counted the total number of words spoken, the number of omitted words beginning with "n", pauses, and grammatical and stylistic errors. All data for the experimental and control groups were calculated and summed, and the obtained results were statistically processed and compared.
Results. The quantitative data of the study, summarized across all indicators, are presented in the table.
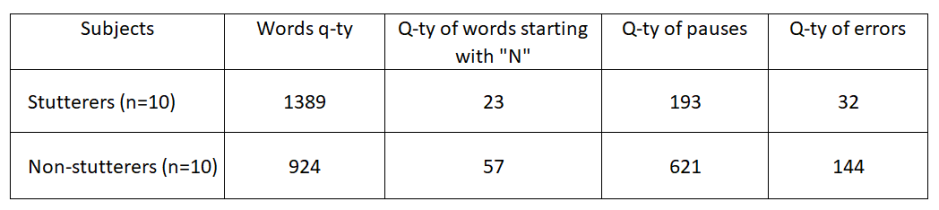
On average, over 3 minutes of retelling, stuttering subjects spoke more words (1389 > 924, p > 0.05). In the speech of stutterers, there were fewer omitted words beginning with "n" compared to representatives of the control group (23 < 57, p > 0.05); fewer pauses between words and sentences (193 < 621, p > 0.05); and fewer grammatical and stylistic errors (32 < 144, p > 0.05). Thus, stutterers handled the task much more easily overall. Their speech during retelling changed only slightly compared to their speech during the preliminary conversation. Non-stuttering subjects retold the text very slowly with many stops and interjections during pauses ("uh", "um", "ah"). Under experimental conditions, the speech of fluent speakers, but not stutterers, showed many grammatical and stylistic errors that were absent during the preliminary conversation. Comparing the nature and ratio of errors in each group led us to conclude: the experimental task overall caused more difficulties for representatives of the control group. It turned out that stutterers cope much more easily with the task of monitoring their own speech and substituting words in a planned utterance than non-stutterers.
Based on an analysis of existing literature data from PET and MRI studies, an impression emerges that stutterers during linguistic activity show activation of the same left-hemisphere structures as non-stutterers, but additionally involve their right-hemisphere homologues in the speech process. The fact that during speech, left-hemisphere sensory and associative areas are more activated in fluent speakers, while right-hemisphere subcortical and frontal areas are more active in stutterers, may mean that stutterers control the speech flow differently from non-stutterers.
Stuttering typically begins in childhood (3-5 years), during the period of active speech development, and leaves a specific imprint on the acquisition of complex linguistic skills. From childhood, the speech activity of a stutterer is aimed simultaneously at forming a competent and coherent utterance and at eliminating the impact of involuntary muscle twitches on expressive speech. Without interrupting the flow of speech, the stutterer must analyze the planned utterance, replace identified "dangerous" words with synonyms, use function words and interjections before disfluencies to reduce muscle contractions, pause during strong spasms, preventing their impact on speech. Possibly, the performance of a complex cognitive-linguistic task of dual speech monitoring is ensured by the additional involvement in linguistic activity of the right-hemisphere homologues of those brain structures that participate in normal linguistic activity. Normally, left-hemisphere Wernicke's area and adjacent areas of the temporal and parietal regions are involved in phonological and semantic preparation of speech [Petersen et al., 1988; Wise et al., 1991; Démonet et al., 1992; Zatorre et al., 1992]. According to Levelt [8] and Belin & Zatorre [2000], activation in the area of the left and right superior temporal gyri is associated not only with the perception of someone else's speech but also with the perception and monitoring of one's own. It is quite possible that the additional involvement of right-hemisphere homologues of brain areas associated with speech activity allows stutterers to successfully cope with the task of dual control over speech and maneuvering between involuntary muscle contractions.
In light of the above, our results may mean that the unusual brain activation in stuttering may reflect, in addition to standard linguistic activity, processes specific to stutterers of monitoring, planning, and restructuring their own speech to avoid manifestations of stuttering and mask their defect. Possibly, the atypical lateralization pattern of linguistic processes observed in stuttering is the result of an individually adapted system that develops from childhood and early adolescence in attempts to speak fluently.
Conclusion. The features of the cerebral organization of speech processes in stutterers may be related to the compensatory-adaptive components of speech and reflect the history of adaptation to their defect and adaptive linguistic behavior.
Literature:
1. Blomgren M., Nagarajan S.S., Lee J.N., Li T., Alvord L. Preliminary results of a functional MRI study of brain activation patterns in stuttering and nonstuttering speakers during a lexical access task. Journal of Fluency Disorders. 2003. V. 28. P. 337--356.
2. Braun A.R., Varga M., Stager S., Schulz G., Selbie S., Maisog J.M., Carson R.E., Ludlow C.L. Altered patterns of cerebral activity during speech and language production in developmental stuttering. An H2(15)O positron emission tomography study. Brain. 1997. V.120. P. 761-784.
3. Chang S.E., Kenney M.K., Loucks T.M., Ludlow C.L. Brain activation abnormalities during speech and non-speech in stuttering speakers. Neuroimage. 2009. V.46. P. 201-212.
4. Ghosh S.S., Tourville J.A., Guenther F.H.. A neuroimaging study of premotor lateralization and cerebellar involvement in the production of phonemes and syllables. J Speech Lang Hear Res. 2008. V. 51(5). P. 1183-1202.
5. Chou T.L., Booth J.R., Bitan T., Burman D.D., Bigio J.D., Cone N.E., Lu D., Cao F. Developmental and skill effects on the neural correlates of semantic processing to visually presented words. Hum Brain Mapp. 2006. V. 27(11). P. 915-924.
6. De Nil L.F., Beal D.S., Lafaille S.J., Kroll R.M., Crawley A.P., Gracco V.L. The effects of simulated stuttering and prolonged speech on the neural activation patterns of stuttering and nonstuttering adults. Brain Lang. 2008. V. 107(2). P. 114-123.
7. Fox P.T., Ingham R.J., Ingham J.C., Zamarripa F., Xiong J.H., Lancaster J.L. Brain correlates of stuttering and syllable production A PET performance-correlation analysis Brain. 2000, V. 123, P. 1985-2004.
8. Levelt W.J., Praamstra P., Meyer A.S., Helenius P., Salmelin R. An MEG study of picture naming. J Cogn Neurosci. 1998. V. 10(5). P. 553-567.
9. Lu C., Chen C., Ning N., Ding G., Guo T., Peng D., Yang Y., Li K., Lin C. The neural substrates for atypical planning and execution of word production in stuttering. Exp Neurol. 2010. V. 221(1). P. 146-56.
10. Seery C.H., Watkins R.V., Mangelsdorf S.C., Shigeto A. Subtyping Stuttering II: Contributions from Language and Temperament. J Fluency Disord. 2007. V. 32(3). P. 197--217.
11. Van Borsel J., Achten E., Santens P., Lahorte P., Voetb T. fMRI of developmental stuttering: A pilot study. Brain and Language. 2003. V. 85. P. 369--376.
12. Watkins K.E., Smith S.M., Davis S., Howell P. Structural and functional abnormalities of the motor system in developmental stuttering. Brain. 2008.131(1). P. 50-59.
Особенности церебральной организации лингвистической деятельности у взрослых заикающихся
Патофизиология заикания до настоящего времени остается объектом изучения современной нейробиологии. За годы исследований были сформулированы многочисленные теории относительно его этиологии и патогенеза. Одной из первых формальных теорий была идея о неполной или аномальной церебральной доминантности, предложенная Orton [1927] и Travis [1931]. Они предположили, что несформированная или нарушенная речевая церебральная доминантность заикающихся приводит к минимизации моторных импульсов к билатерально иннервируемым речевым мышцам. С 1966 года, когда Jones, используя тест Wada, установил билатеральное представительство речи у 4 заикающихся, появились новые свидетельства аномального паттерна латерализации у данной категории больных. Основная масса результатов подтверждала, что заикающиеся демонстрируют различия в церебральной доминантности по сравнению с нормой [Fox и др., 1996; Ingham и др., 1994; Ingham и др., 1996; Wu и др., 1995]. По результатам этих работ были высказаны разные предположения о локализации речи: левополушарная доминантность, правополушарная доминантность, вмешательство правого полушария в работу левого, инверсия речевой доминантности, билатеральное представительство речи.
В последние годы позитронно-эмиссионная (ПЭТ) и магнитно-резонансная томография (МРТ) открыли новые возможности для исследования морфофункционального субстрата речи, как в норме, так и при заикании. Эти исследования уточнили существующие представления о локализации сенсомоторных и когнитивно-лингвистических компонентов речи. В норме запланированная и подготовленная речь без семантической составляющей активирует первичную моторную зону - 4 поле (здесь и далее поля по Brodmann) билатерально [3, 5, 7, 11]. Правая и левая первичные моторные области симметрично активируются, поскольку артикуляционные, дыхательные мышцы, а также глотка и гортань иннервируются билатерально. Только слева при реализации речи активируется вторичная моторная область коры (6) [2, 4, 5]. Региональный мозговой кровоток в дорсолатеральных отделах префронтальной коры (8, 9), медиальной орбитальной (11) и медиальной префронтальной (10) областях, предположительно связанных с семантическими компонентами речевой деятельности, в норме повышается только слева [1, 2]. Области первичной слуховой и слуховой ассоциативной коры в верхней височной извилине (41, 42, 22) в норме активируются билатерально. После первичной и вторичной обработки в этих областях информация передается во фронтальные, теменно-височные, паралимбические и подкорковые области [Pandya&Yeterian, 1985]. Таким образом, эти участки коры связаны не только со слуховой перцепцией, но и с подготовкой экспрессивной речи [2, 8, 11]. Верхние оперкулярные области (44, 45), классически составляющие поле Broca, традиционно считаются участвующими в речевом моторном программировании [Freedman и др., 1984] и демонстрируют унилатеральную левостороннюю активацию в норме. Передневентральные области фронтальной покрышки (47), активируемые слева, более тесно связаны с синтаксическими процессами [McCarthy и др., 1993] и формированием высказывания [1, 4, 11]. Только в левом полушарии в норме при лингвистических процессах активируются передний височный полюс (38) и угловая извилина (39) [2, 11]. John Van Borsel с соавторами обнаружили включение в процесс речи третичных полей – моторных (46 билатерально), зрительных (19 слева, 7 справа) и сенсорных (40 билатерально) [11].
Помимо корковых областей в обеспечении речи у здоровых испытуемых правшей участвует и мозжечок. Считается, что это образование связано со слуховой корой и содержит нейроны, которые отвечают за слуховые стимулы [Snider и др., Stowell, 1944; Hampson 1949; Aitkin&Boyd, 1975; Huang&Liu, 1985]. Vermis мозжечка располагается в позиции, позволяющей контролировать речевую продукцию и через восходящие эфференты модулировать речевую моторную активность. Таким образом, мозжечок принимает участие в контроле над речью и специфически вовлечен в выявление происходящих ошибок и коррекцию моторной активности, инициированной неокортикальными системами [Thach и др., 1992].
Insula в норме активируется акустической стимуляцией [Kushner и др., 1987], и избирательное поражение этой области может привести к относительно специфичному нарушению обработки слуховых сигналов [Fifer, 1993]. Mohr в 70-х провел тщательное сравнительное клиническое и посмертное изучение мозга больных и установил, что инфаркты, ограниченные областью левой передней insula и pars opercularis приводят к временной тяжелейшей дизартрии или почти немоте без сопутствующей афазии [Mohr и др., 1978]. По некоторым данным, insula может быть вовлечена в относительно прямую обработку слуховой информации, по другим - в речевое артикуляторное планирование [Pandya и др., 1969; Mesulam&Mufson, 1982].
Согласно ПЭТ–исследованиям, метаболизм мозга заикающихся и незаикающихся в состоянии покоя не различается [Braun и др., 1997; Ingham и др., 1996]. Артикуляционные и глоточные движения при реализации речи без лингвистической составляющей вызывают билатеральную активацию в первичной моторной, премоторной, соматосенсорной и слуховой коре, островке, мозжечке, скорлупе, вентральном таламусе, как в норме, так и при заикании. В то время как качественные различия в природе и анатомическом распределении активации отсутствуют, магнитуда увеличения регионального кровотока в этих областях во время реализации речи значительно больше у заикающихся [2, 11]. В неокортексе активация больше в премоторной, первичной моторной и соматосенсорной коре, что означает, что эти области могут быть более активны в генерации и проприоцептивной или тактильной перцепции движений губ, языка, челюсти и гортани. Многие исследователи получили доказательства сверхактивности правой первичной моторной зоны (4) заикающихся во время речи по сравнению с нормой [2, 11, 12]. Медиальная часть поля 6 дополнительной моторной коры, связанная предположительно с речевым дыханием, ограничена левым полушарием и в норме и при заикании, тогда как латеральная часть поля 6, связанная, возможно, с моторным и фонематическим планированием, при заикании активируется билатерально со сниженной активацией слева [2, 4, 5].
Церебральная организация лингвистических компонентов речи, в отличие от сенсомоторных, фундаментально отличается у взрослых с заиканием. В норме во время формулирования речевого высказывания мозговой кровоток главным образом повышается в левом полушарии. У заикающихся повышение кровотока отсутствует, пространственно ограничено, билатерально или латерализовано в правом полушарии [2, 3, 6, 9, 11, 12]. Вторичные двигательные (8, 9), медиальные орбитальная (11) и префронтальная (10) области демонстрируют унилатеральную левостороннюю активацию в норме и слабую билатеральную активацию у заикающихся [Chang и др., 2009; Wood и др., 1980; Wu и др., 1995; Fox и др., 2000]. Первичные и вторичные слуховые поля (41, 42, 22), в норме активируются билатерально, а у заикающихся кровоток слева в первичной слуховой и слуховой ассоциативной коре значительно ниже нормы, даже в состоянии покоя [Pool и др., 1991]; зато правосторонний гомолог области Wernicke (22) более активен [3, 6, 7, 9, 12]. Передний височный полюс (38), активируемый при лингвистических процессах в норме слева, активируется у заикающихся справа. Кроме того, заикающиеся не демонстрируют активацию левых височно-теменных областей, которые граничат с зоной Wernicke – задней верхней височной (40) и нижней угловой извилин (39) [2, 11]. Только у заикающихся наблюдается активация правого гомолога области Broca (44, 45) [1, 2].
Во время речи у заикающихся значительно повышен кровоток в мозжечке билатерально, в insula билатерально с распространением на putamen, и среднем мозге (substantia nigra, pedunculopontine nucleus, red nucleus и subthalamic nucleus) билатерально [12]. Только у заикающихся активируется anterior insula справа, putamen справа и мезенцефалическое околоводопроводное серое вещество [2, 3, 6, 7, 9, 11].
До сегодняшнего дня не выявлено, является ли специфический паттерн активации мозга при заикании причиной дефекта, отражением компенсаторной деятельности нервной системы или следствием приспособительного поведения.
Взрослые заикающиеся сообщают, что они с детского или подросткового возраста контролируют свою речь для того, чтобы замаскировать проявления дефекта и попытаться предотвратить возникающие запинки с помощью различных приемов и уловок. К наиболее распространенным речевым уловкам относятся: замена слов на те, в которых не встречались бы определенные «трудные» звуки; периодические остановки внутри слов и между словами для того, чтобы исключить влияние непроизвольных сокращений артикуляционных, вокальных и глоточных мышц на речь; частое использование перед трудными словами стандартных, привычных и легких служебных слов, не несущих семантической нагрузки («ну», «это», «вот» и др.). По видимому, заикающемуся во время речи приходится предварительно анализировать предстоящее высказывание и, если в нем встречаются слова, начинающиеся с проблемных для него звуков, предварять их служебными словами или заменять на синонимы; останавливаться во время непроизвольных мышечных сокращений. При этом лингвистическая деятельность заикающегося с детства должна быть направлена не только на построение грамотного и распространенного речевого высказывания, наиболее точно отражающего его мысли и чувства, но и на то, чтобы минимизировать проявления дефекта в речи. Если предположить, что лингвистические процессы заикающегося направлены одновременно и на формирование предстоящего высказывания и на сканирование будущего предложения, то, в таком случае, заикающиеся, по сравнению с плавноговорящими людьми, должны были бы лучше выполнять речевые задачи, связанные с избеганием определенных слов и звуков. Чтобы проверить это предположение, мы предложили 2 группам испытуемых рассказывать хорошо знакомый текст, избегая слов, начинающихся с определенных букв.
Методы. В исследовании принимали участие 10 заикающихся и 10 незаикающихся правшей. Гендерный состав (8м/2ж), средний возраст (45±9 лет) и уровень образования (12±4 года обучения) были в среднем равными в обеих группах. Каждый из испытуемых должен был прочитать текст, состоящий из 1000 слов. После этого испытуемому предлагали пересказать его, не используя слова, начинающиеся с буквы «н» (в тексте было 45 слов, начинающихся с буквы "н"). В течение 3 минут пересказа экспериментатор подсчитывал общее количество произнесенных слов, количество пропущенных слов на букву «н», остановок, грамматических и стилистических ошибок. Все данные по экспериментальной и контрольной группам подсчитывали и суммировали, полученные результаты статистически обрабатывали и сравнивали.
Результаты. Количественные данные исследования, суммированные по всем показателям, приведены в таблице.
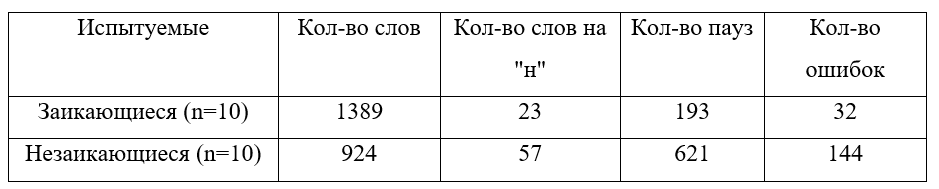
В среднем за 3 минуты пересказа заикающиеся испытуемые произнесли больше слов (1389>924, p>0,05). В речи заикающихся оказалось меньше пропущенных слов, начинающихся с «н» по сравнению с представителями контрольной группы (23<57, p>0,05); меньше пауз между словами и предложениями (193<621, p>0,05); меньше грамматических и стилистических ошибок (32<144, p>0,05). Таким образом, заикающиеся в целом гораздо легче справились с поставленной задачей. Их речь во время пересказа незначительно изменилась по сравнению с речью во время предварительной беседы. Незаикающиеся испытуемые пересказывали текст очень медленно с большим количеством остановок и междометий во время пауз ("эээ", "ммм", "ааа""). В условиях эксперимента в речи плавноговорящих людей, но не заикающихся, появилось большое количество грамматических и стилистических ошибок, отсутствующих во время предварительной беседы. Сравнение характера и соотношения ошибок в каждой группе привело нас к выводу: в целом экспериментальная задача вызвала больше затруднений у представителей контрольной группы. Оказалось, что заикающиеся гораздо легче справляются с задачей мониторирования собственной речи и замене слов в планируемом высказывании, чем незаикающиеся.
На основании анализа существующих литературных данных ПЭТ и МРТ исследований складывается представление, что заикающиеся в процессе реализации лингвистической деятельности показывают активацию тех же структур слева, что и незаикающиеся, но дополнительно вовлекают их правосторонние гомологи в процесс речи. Тот факт, что во время речи левополушарные сенсорные и ассоциативные области более активированы у плавноговорящих людей, тогда как правополушарные подкорковые и фронтальные области в большей степени активны у заикающихся, может означать, что заикающиеся контролируют речевой поток иначе, чем незаикающиеся.
Заикание, как правило, начинается в детском возрасте (3-5 лет), в период активного развития речи и специфическим образом накладывает отпечаток на приобретение сложных лингвистических навыков. С детских лет речевая деятельность заикающегося направлена одновременно и на формирование грамотного и согласованного высказывания, и на то, чтобы элиминировать воздействие непроизвольных мышечных подергиваний на экспрессивную речь. Не прерывая потока речи, заикающийся должен анализировать запланированное высказывание, заменять выявленные «опасные» слова на синонимы, использовать служебные слова и междометия перед запинками для ослабления мышечных сокращений, делать паузы во время сильных судорог, не допуская их воздействия на речь. Возможно, выполнение сложной когнитивно-лингвистической задачи по двойному мониторированию речи обеспечивается дополнительным вовлечением в лингвистическую активность правосторонних гомологов тех структур мозга, которые участвуют в нормальной лингвистической деятельности. В норме в фонологическую и семантическую подготовку речи вовлечены левополушарная область Wernicke и прилегающие к ней участки височной и теменной областей [Petersen и др., 1988; Wise и др., 1991; De´monet и др., 1992; Zatorre и др., 1992]. Согласно данным Levelt [8] и Belin&Zatorre [2000], активация в области левой и правой верхней височной извилин связана не только с восприятием чужой речи, но и с перцепцией и мониторированием собственной. Вполне возможно, что дополнительное вовлечение правосторонних гомологов областей мозга, связанных с речевой деятельностью, позволяет заикающимся успешно справляться с задачей двойного контроля над речью и лавирования между непроизвольными мышечными сокращениями.
В свете вышесказанного наши результаты могут означать, что необычная активация мозга при заикании может отражать, помимо стандартной лингвистической деятельности, и специфические для заикающихся процессы мониторирования, планирования и перестройки собственной речи с целью избегания проявлений заикания и маскировки своего дефекта.. Возможно, атипичный паттерн латерализации лингвистических процессов, наблюдаемый при заикании, является результатом индивидуально адаптированной системы, которая развивается с детского и раннего подросткового возраста в попытках говорить плавно.
Заключение. Особенности церебральной организации речевых процессов заикающихся могут быть связаны с компенсаторно-приспособительными составляющими речи и отражают историю приспособления к своему дефекту и адаптивное лингвистическое поведение.
Литература:
1. Blomgren M., Nagarajan S.S., Lee J.N., Li T., Alvord L. Preliminary results of a functional MRI study of brain activation patterns in stuttering and nonstuttering speakers during a lexical access task. Journal of Fluency Disorders. 2003. V. 28. P. 337-356.
2. Braun A.R., Varga M., Stager S., Schulz G., Selbie S., Maisog J.M., Carson R.E., Ludlow C.L. Altered patterns of cerebral activity during speech and language production in developmental stuttering. An H2(15)O positron emission tomography study. Brain. 1997. V.120. P. 761-784.
3. Chang S.E., Kenney M.K., Loucks T.M., Ludlow C.L. Brain activation abnormalities during speech and non-speech in stuttering speakers. Neuroimage. 2009. V.46. P. 201-212.
4. Ghosh S.S., Tourville J.A., Guenther F.H.. A neuroimaging study of premotor lateralization and cerebellar involvement in the production of phonemes and syllables. J Speech Lang Hear Res. 2008. V. 51(5). P. 1183-1202.
5. Chou T.L., Booth J.R., Bitan T., Burman D.D., Bigio J.D., Cone N.E., Lu D., Cao F. Developmental and skill effects on the neural correlates of semantic processing to visually presented words. Hum Brain Mapp. 2006. V. 27(11). P. 915-924.
6. De Nil L.F., Beal D.S., Lafaille S.J., Kroll R.M., Crawley A.P., Gracco V.L. The effects of simulated stuttering and prolonged speech on the neural activation patterns of stuttering and nonstuttering adults. Brain Lang. 2008. V. 107(2). P. 114-123.
7. Fox P.T., Ingham R.J., Ingham J.C., Zamarripa F., Xiong J.H., Lancaster J.L. Brain correlates of stuttering and syllable production A PET performance-correlation analysis Brain. 2000, V. 123, P. 1985-2004.
8. Levelt W.J., Praamstra P., Meyer A.S., Helenius P., Salmelin R. An MEG study of picture naming. J Cogn Neurosci. 1998. V. 10(5). P. 553-567.
9. Lu C., Chen C., Ning N., Ding G., Guo T., Peng D., Yang Y., Li K., Lin C. The neural substrates for atypical planning and execution of word production in stuttering. Exp Neurol. 2010. V. 221(1). P. 146-56.
10. Seery C.H., Watkins R.V., Mangelsdorf S.C., Shigeto A. Subtyping Stuttering II: Contributions from Language and Temperament. J Fluency Disord. 2007. V. 32(3). P. 197--217.
11. Van Borsel J., Achten E., Santens P., Lahorte P., Voetb T. fMRI of developmental stuttering: A pilot study. Brain and Language. 2003. V. 85. P. 369--376.
12. Watkins K.E., Smith S.M., Davis S., Howell P. Structural and functional abnormalities of the motor system in developmental stuttering. Brain. 2008.131(1). P. 50-59.